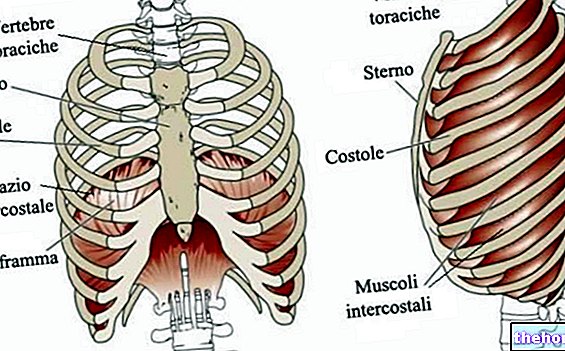
A fim de ser capaz de falar de uma forma compreensível do "hemoglobina (Hb), é útil cuidar do primeiro mioglobina (Mb) que é muito semelhante à hemoglobina, mas é muito mais simples. Entre a hemoglobina e a mioglobina existem relações de parentesco próximas: ambas são proteínas conjugadas e seu grupo protético (parte não proteica) é o grupo heme.
A mioglobina é uma proteína globular que consiste em uma única cadeia de cerca de cento e cinquenta aminoácidos (depende do organismo) e seu peso molecular é de cerca de 18 Kd.
Como mencionado, ele é equipado com um grupo heme que é inserido em uma porção hidrofóbica (ou lipofílica) da proteína, consistindo em dobras atribuíveis às estruturas em hélice α das proteínas fibrosas.
A mioglobina é composta principalmente por segmentos de hélices α, presentes em número de oito e consiste, quase exclusivamente, em resíduos apolares (leucina, valina, metionina e fenilalanina) enquanto os resíduos polares estão praticamente ausentes (ácido aspártico, ácido glutâmico, lisina e arginina); os únicos resíduos polares são duas histidinas, que desempenham um papel fundamental na ligação do oxigênio ao grupo heme.
O grupo heme é um grupo cromóforo (absorve no visível) e é o grupo funcional da mioglobina.
Veja também: hemoglobina glicada - hemoglobina na urina
Um pouco de quimica

A ligação entre a protoporfirina e o ferro é uma ligação típica de compostos de coordenação que são compostos químicos nos quais um átomo central (ou íon) forma ligações com outras espécies químicas em um número maior que seu número de oxidação (carga elétrica). No caso do heme, essas ligações são reversíveis e fracas.
O número de coordenação (número de ligações de coordenação) do ferro é seis: pode haver seis moléculas ao redor do ferro compartilhando os elétrons de ligação.
Para formar um composto de coordenação, são necessários dois orbitais com orientação correta: um capaz de "adquirir" elétrons e outro capaz de doá-los.
No heme, o ferro forma quatro ligações planas com os quatro átomos de nitrogênio no centro do anel de proto-porfirina e uma quinta ligação com o nitrogênio proximal da histidina; o ferro tem a sexta ligação de coordenação livre e pode se ligar ao oxigênio.
Quando o ferro está na forma de um íon livre, seu tipo orbitais d todos eles têm a mesma energia; na mioglobina, o íon ferro está ligado à protoporfirina e histidina: essas espécies perturbam magneticamente os orbitais d um pouco de ferro; a extensão da perturbação será diferente para os vários orbitais d dependendo de sua orientação espacial e das espécies perturbadoras. Como a energia total dos orbitais deve ser constante, a perturbação provoca uma separação energética entre os vários orbitais: a energia adquirida por alguns orbitais é equivalente à energia perdida por outros.
Se a separação que ocorre entre os orbitais não for muito grande, um arranjo eletrônico de spin alto é preferível: os elétrons de ligação tentam se organizar em spins paralelos em tantos subníveis quanto possível (multiplicidade máxima); se, por outro lado, a perturbação for muito forte e houver uma grande separação entre os orbitais, pode ser mais conveniente emparelhar os elétrons de ligação nos orbitais de baixa energia (spin baixo).
Quando o ferro se liga ao oxigênio, a molécula assume um arranjo de spin baixo, enquanto quando o ferro tem a sexta ligação de coordenação livre, a molécula tem um arranjo de spin alto.
Graças a essa diferença de spin, por meio de uma análise espectral da mioglobina, podemos entender se o oxigênio (MbO2) está ligado a ela ou não (Mb).
A mioglobina é uma proteína muscular típica (mas não é encontrada apenas nos músculos).
A mioglobina é extraída do cachalote no qual está presente em grandes quantidades e é então purificada.
Os cetáceos têm uma respiração semelhante à dos humanos: tendo pulmões, devem absorver o ar pelo processo respiratório; o cachalote deve trazer o máximo de oxigênio possível para os músculos que são capazes de acumular oxigênio ligando-se à mioglobina presente neles; o oxigênio é então liberado lentamente quando o cetáceo é imerso porque seu metabolismo requer oxigênio: quanto maior a quantidade de oxigênio que o cachalote é capaz de absorver e mais oxigênio fica disponível durante o mergulho.
A mioglibina se liga ao oxigênio de maneira reversível e está presente nos tecidos periféricos em maior porcentagem quanto mais esse tecido está acostumado a trabalhar com suprimentos de oxigênio distantes no tempo.
<--- A mioglobina é uma proteína presente nos músculos, cuja função é justamente a de "reservatório" de oxigênio.

O que torna a carne mais ou menos vermelha é o conteúdo de hemoproteínas (é o heme que torna a carne vermelha).
A hemoglobina tem muitas semelhanças estruturais com a mioglobina e é capaz de se ligar ao oxigênio molecular de maneira reversível; mas, enquanto a mioglobina está confinada aos músculos e tecidos periféricos em geral, a hemoglobina é encontrada nos eritrócitos ou nas células vermelhas do sangue (são pseudocélulas, ou seja, não são células reais) que constituem 40% do sangue.
Ao contrário da mioglobina, o trabalho da hemoglobina é levar oxigênio aos pulmões, liberá-lo nas células onde é necessário, pegar dióxido de carbono e liberá-lo nos pulmões, onde o ciclo começa novamente.
EU"hemoglobina é um tetrâmetro, ou seja, é composto por quatro cadeias polipeptídicas cada uma com um grupo heme e idênticas dois a dois (no ser humano existem duas cadeias alfa e duas cadeias beta).
A principal função da hemoglobina é o transporte de oxigênio; outra função do sangue na qual a hemoglobina está envolvida é o transporte de substâncias para os tecidos.
No trajeto dos pulmões (ricos em oxigênio) aos tecidos, a hemoglobina carrega oxigênio (ao mesmo tempo que as demais substâncias chegam aos tecidos) enquanto no caminho inverso carrega consigo os resíduos coletados pelos tecidos, principalmente o carbono dióxido produzido no metabolismo.
No desenvolvimento de um ser humano existem genes que se expressam apenas por um determinado período de tempo; por isso existem diferentes hemoglobinas: fetais, embrionárias, do homem adulto.
As cadeias que compõem essas diferentes hemoglobinas têm estruturas diferentes, mas com algumas semelhanças, na verdade, a função que desempenham é mais ou menos a mesma.
Uma explicação para a presença de várias cadeias diferentes é a seguinte: no curso do processo evolutivo dos organismos, até a hemoglobina evoluiu especializando-se no transporte de oxigênio de áreas ricas nele para áreas deficientes. da cadeia evolutiva l "hemoglobina transportou oxigênio em pequenos organismos; no curso da evolução os organismos alcançaram dimensões maiores, portanto a hemoglobina foi modificada para poder transportar oxigênio para áreas mais distantes do ponto em que era rico nele; para para isso, foram codificadas, no decorrer do processo evolutivo, novas estruturas das cadeias que constituem a hemoglobina.
A mioglobina se liga ao oxigênio mesmo em pressões modestas; nos tecidos periféricos há uma pressão (PO2) de cerca de 30 mmHg: a mioglobina nessa pressão não libera oxigênio, por isso seria ineficaz como transportadora de oxigênio. A hemoglobina, por outro lado , tem um comportamento mais elástico: liga o oxigênio a altas pressões e o libera quando a pressão diminui.
Quando uma proteína é funcionalmente ativa, ela pode mudar um pouco de forma; por exemplo, a mioglobina oxigenada tem uma forma diferente da mioglobina não oxigenada e essa mutação não afeta seus vizinhos.
A situação é diferente no caso de proteínas associadas, como a hemoglobina: quando uma cadeia se oxigena é induzida a mudar de forma, mas essa modificação é tridimensional, de modo que as outras cadeias do tetrâmetro também são afetadas. O fato de as cadeias estarem associadas entre si., sugere que a modificação de um afeta os outros vizinhos, mesmo que em uma extensão diferente; quando uma cadeia se oxigena, as outras cadeias do tetrâmetro assumem uma "atitude menos hostil" em relação ao oxigênio: a dificuldade com que uma cadeia é oxigenado diminui à medida que as cadeias próximas a ele oxigenam, por sua vez. O mesmo vale para a desoxigenação.
A estrutura quaternária da desoxiemoglobina é chamada de forma T (tensa), enquanto a da oxiemoglobina é chamada de forma R (liberada); no estado tenso, há uma série de interações eletrostáticas bastante fortes entre os aminoácidos ácidos e os aminoácidos básicos que levam a uma estrutura rígida da desoxiemoglobina (é por isso que a "forma tensa"), enquanto quando o oxigênio está ligado, a entidade destes as interações diminuem (daí a "forma liberada"). Além disso, na ausência de oxigênio, a carga da histidina (ver estrutura) é estabilizada pela carga oposta do ácido aspártico enquanto, na presença de oxigênio, há uma tendência por parte da proteína de perder um próton; tudo isso envolve que a hemoglobina oxigenada é um ácido mais forte do que a hemoglobina desoxigenada: efeito bohr.
Dependendo do pH, o grupo heme se liga mais ou menos facilmente ao oxigênio: em um ambiente ácido, a hemoglobina libera oxigênio mais facilmente (a forma tensa é estável) enquanto, em um ambiente básico, a ligação com o oxigênio é mais difícil.

Cada hemoglobina libera 0,7 prótons por mol de oxigênio (O2) que entra.
O efeito Bohr permite que a hemoglobina melhore sua capacidade de transportar oxigênio.
A hemoglobina que viaja dos pulmões para os tecidos deve se equilibrar em função da pressão, pH e temperatura.
Vamos ver o efeito da temperatura.
A temperatura nos alvéolos pulmonares é cerca de 1-1,5 ° C mais baixa do que a temperatura externa, enquanto nos músculos a temperatura é cerca de 36,5-37 ° C; com o aumento da temperatura, o fator de saturação diminui (na mesma pressão): isso ocorre porque a energia cinética aumenta e a dissociação é favorecida.

Existem outros fatores que podem afetar a capacidade da hemoglobina de se ligar ao oxigênio, um dos quais é a concentração de 2,3 bisfosfoglicerato.
O 2,3 bisfosfoglicerato é um metabólico presente nos eritrócitos em uma concentração de 4-5 mM (em nenhuma outra parte do organismo está presente em uma concentração tão alta).
Em pH fisiológico, o 2,3 bisfosfoglicerato é desprotonado e possui cinco cargas negativas; ele está preso entre as duas cadeias beta da hemoglobina porque essas cadeias têm uma alta concentração de cargas positivas. As interações eletrostáticas entre as cadeias beta e o 2,3 bisfosfoglicerato conferem certa rigidez ao sistema: obtém-se uma estrutura tensa com pouca afinidade pelo oxigênio; durante a oxigenação, o 2,3 bisfosfoglicerato é expelido.
Nos eritrócitos, c "é um aparato especial que converte o 1,3 bisfosfoglicerato (produzido pelo metabolismo) em 2,3 bisfosfoglicerato de forma que atinge uma concentração de 4-5 mM e, portanto, a hemoglobina é capaz de trocar o" oxigênio nos tecidos.
A hemoglobina que chega ao tecido está no estado liberado (ligada ao oxigênio), mas na vizinhança do tecido, ela é carboxilada e passa ao estado tenso: a proteína neste estado tem menos tendência a se ligar ao oxigênio, com respeito para o estado liberado, portanto, a hemoglobina libera oxigênio para o tecido; além disso, por reação entre a água e o dióxido de carbono, ocorre a produção de íons H +, portanto mais oxigênio devido ao efeito bohr.
O dióxido de carbono se difunde para o eritrócito, passando pela membrana plasmática; uma vez que os eritrócitos constituem cerca de 40% do sangue, devemos esperar que apenas 40% do dióxido de carbono que se difunde dos tecidos entre neles; na verdade, 90% do dióxido de carbono entra nos eritrócitos porque contêm uma enzima que converte o dióxido de carbono no ácido carbônico, resulta que a concentração estacionária de dióxido de carbono nos eritrócitos é baixa e, portanto, a taxa de entrada é alta.

Outro fenômeno que ocorre quando um eritrócito atinge um tecido é o seguinte: por gradiente, o "HCO3- (derivado do dióxido de carbono) sai do" eritrócito e, para equilibrar a saída de uma carga negativa, temos a "entrada de cloretos que determina um aumento da pressão osmótica: para equilibrar esta variação há também a entrada de água que causa inchaço do eritrócito (efeito HAMBURGER). O fenômeno oposto ocorre quando um eritrócito atinge os alvéolos pulmonares: uma desinsuflação dos eritrócitos (efeito HALDANE) Portanto, os eritrócitos venosos (direcionados aos pulmões) são mais arredondados que os arteriais.