Editado pelo Dr. Giovanni Chetta
ECM é geralmente descrito como sendo composto de várias classes grandes de biomoléculas:
- Proteínas estruturais (colágeno e elastina)
- Proteínas especializadas (fibrilina, fibronectina, laminina etc.)
- Proteoglicanos (agrecanos, sindecanos) e glusaminoglicanos (hialuronanos, sulfatos de condroitina, sulfatos de heparano, etc.)
Proteínas estruturais
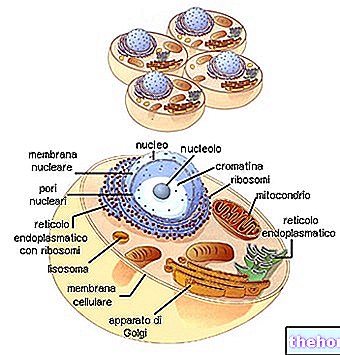
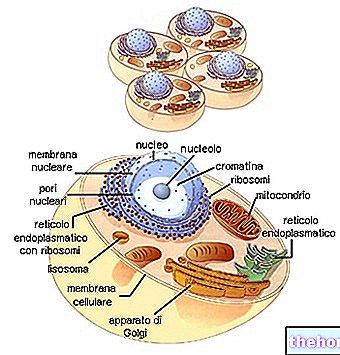
Os colágenos formam a família de glicoproteínas mais representada no reino animal. São as proteínas mais presentes na matriz extracelular (mas não as mais importantes) e são os constituintes fundamentais dos próprios tecidos conjuntivos (cartilagem, osso, fáscia, tendões, ligamentos).
Existem pelo menos 16 tipos diferentes de colágeno, dos quais os tipos I, II e III são os mais presentes ao nível das fibrilas típicas (o tipo IV forma uma espécie de retículo que representa o principal componente das lâminas basais).
Os colágenos são principalmente sintetizados por fibroblastos, mas as células epiteliais também são capazes de sintetizá-los.
As fibras de colágeno interagem continuamente com uma enorme quantidade de outras moléculas da matriz extracelular, constituindo um continuum biológico fundamental para a vida da célula. Os colágenos associados às fibrilas desempenham um papel predominante na formação e manutenção de estruturas capazes de resistir às forças de tensão, sendo quase inelástico (os glicosamlicanos desempenham uma ação de resistência à compressão). De alguma forma o colágeno é produzido e remetabolizado em função da carga mecânica e suas propriedades viscoelásticas acarretam, como veremos no parágrafo "Viscoelasticidade de a fáscia ", um grande impacto na postura do homem. Como mais uma demonstração da capacidade do colágeno de mudar de acordo com as influências ambientais, assumindo, por exemplo, Em graus variáveis de rigidez, elasticidade e resistência, existem colágenos, definidos com o termo FACIT (Colágeno Associado a Fibrilas com Hélices Triplas Interrompidas) capazes de funcionar funcionalmente como proteoglicanos (descritos no parágrafo “Glucosaminoglicanos e proteoglicanos”).
As fibras de colágeno, graças ao seu revestimento de PG / GAG (proteoglicanos / glucosaminoglicanos) possuem propriedades de biossensores e biocondutores: as cargas elétricas relativas resultam em uma maior capacidade de ligar água e trocar íons, portanto, uma maior capacidade elétrica.
Sabemos que qualquer força mecânica capaz de gerar uma deformação estrutural tensiona as ligações inter-moleculares, produzindo um leve fluxo elétrico, que é o corrente piezoelétrica (Athenstaedt, 1969). Nesses casos, as fibras de colágeno distribuem as cargas positivas em sua superfície convexa e as negativas na côncava, transformando-se em semicondutores (permitem o fluxo de elétrons em sua superfície unidirecional). Uma vez que a energia piezoelétrica (bem como a energia piroelétrica gerada por tensões térmicas) é neutralizada pelos íons circulantes em um tempo muito curto (aprox. 10-7 - 10-9 segundos), o arranjo do PG / GAG no sinal é decisivo para a propagação do sinal na superfície das fibrilas, de forma a atuar como "repetidores" do impulso elétrico. Em particular, uma periodicidade longitudinal de aprox. 64 nm (que ao microscópio óptico aparece como uma linha) permite uma velocidade de propagação do impulso igual a cerca de 64 m / s (correspondendo à velocidade de condução das fibras nervosas rápidas) - Rengling, 2001. O forte momento dipolar das fibrilas de colágeno e sua capacidade de ressonância (propriedade comum a todas as estruturas peptídicas), bem como a baixa constante dielétrica do MEC, facilitam a transmissão de sinais eletromagnéticos. Portanto, a rede tridimensional e ubíqua de colágeno também possui a característica peculiar de conduzir sinais bioelétricos em as três dimensões do espaço, baseadas no arranjo relativo entre as fibrilas e células de colágeno, na direção aferente (da MEC para as células) ou, vice-versa, eferente.
Tudo isso representa um sistema de comunicação de células MEC em tempo real e tais biossinais eletromagnéticos podem levar a alterações bioquímicas importantes, por exemplo, no osso, os osteoclastos não podem "digerir" ossos com carga piezoelétrica (Oschman, 2000).
Por fim, deve-se ressaltar que a célula, não surpreendentemente, produz continuamente e com considerável dispêndio de energia (aprox. 70%) material que deve necessariamente ser expelido, principalmente através do armazenamento exclusivo de protocolágeno (precursor biológico do colágeno) em específico vesículas (Albergati, 2004).
A grande maioria dos tecidos de vertebrados requer a presença simultânea de duas características vitais: força e elasticidade. Uma verdadeira rede de fibras elásticas, localizada dentro da MEC desses tecidos, permite o retorno às condições iniciais após fortes trações, podendo aumentar a extensibilidade de um órgão ou de parte dele em pelo menos cinco vezes. Fibras de colágeno longas e inelásticas são intercaladas entre as fibras elásticas com a tarefa precisa de limitar a "deformação excessiva devido à tração dos tecidos. L"elastina representa o principal componente das fibras elásticas. É uma proteína extremamente hidrofóbica, com cerca de 750 aminoácidos de comprimento, pois o colágeno é rico em prolina e glicina mas, ao contrário do colágeno, não é glicado e contém muitos resíduos de hidroxiprolina e não hidroxilisina. A elastina surge como uma verdadeira rede bioquímica de forma irregularmente tridimensional, composta por fibras e lamelas que permeiam a MEC de todos os tecidos conjuntivos. Encontra-se em quantidades particularmente abundantes nos vasos sanguíneos com características elásticas (é a proteína da MEC mais presente nas artérias e representa mais de 50% do peso seco total da aorta), nos ligamentos, no pulmão e na pele. Na derme, ao contrário do que acontece com o colágeno, a densidade e o volume da elastina tendem a aumentar com o tempo, mas a elastina velha geralmente aparece inchada, quase inchada, muitas vezes com aspecto fragmentado e com redução do componente "amorfo". (Pasquali Rochetti et al, 2004). As células musculares lisas e os fibroblastos são os principais produtores de seu precursor, a tropoelastina, secretada nos espaços extracelulares.
Outros artigos sobre "Colágeno e elastina, fibras de colágeno na matriz extracelular"
- Matriz extracelular
- Fibronectina, Glucosaminoglicanos e Proteoglicanos
- Importância da matriz extracelular no equilíbrio celular
- Alterações da matriz extracelular e patologias
- Tecido conjuntivo e matriz extracelular
- Fáscia profunda - tecido conjuntivo
- Mecanorreceptores fasciais e miofibroblastos
- Biomecânica da fáscia profunda
- Postura e equilíbrio dinâmico
- Tensegridade e movimentos helicoidais
- Membros inferiores e movimento corporal
- Suporte da culatra e aparelho estomatognático
- Casos clínicos, alterações posturais
- Casos clínicos, postura
- Avaliação postural - caso clínico
- Bibliografia - Da matriz extracelular à postura. O sistema conectivo é nosso verdadeiro Deus ex machina?