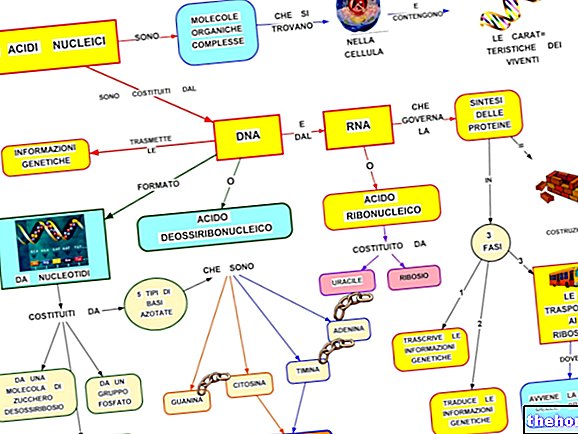
Para que haja correspondência entre a informação do polinucleotídeo e a do polipeptídeo, existe um código: o código genético.
As características gerais do código genético podem ser listadas da seguinte forma:
O código genético é composto de trigêmeos e não contém pontuação interna (Crick & Brenner,).
Ele "foi decifrado através do uso de" sistemas de tradução de células abertas "(Nirenberg & Matthaei, 1961; Nirenberg & Leder, 1964; Korana, 1964).
É altamente degenerado (sinônimos).
A organização da tabela de códigos não é acidental.
Trigêmeos "absurdos".
O código genético é "padrão", mas não "universal".
Olhando para a tabela do código genético, deve ser lembrado que se refere à tradução de "RNAm em polipeptídeo, para o qual as bases de nucleotídeos envolvidas são A, U, G, C. A biossíntese de uma cadeia polipeptídica é a tradução de a sequência de nucleotídeos na sequência de aminoácidos.

Cada tripleto de base do RNAm, denominado códon, tem a primeira base na coluna da esquerda, a segunda na linha superior e a terceira na coluna da direita. Tomemos, por exemplo, o triptofano (ou seja, Try) e veremos que o códon correspondente seja, em ordem, UGG. Na verdade, a primeira base, U, inclui toda a linha de caixas no topo; neste, G identifica a caixa mais à direita e a quarta linha da própria caixa, onde encontramos a escrita Try. Da mesma forma, para sintetizar o tetrapeptídeo Leucina-Alanina-Arginina-Serína (símbolos Leu-Ala-Arg-Ser), podemos encontrar os códons UUA-AUC-AGA-UCA no código.

Neste ponto, entretanto, deve-se notar que todos os aminoácidos de nosso tetrapeptídeo são codificados (ao contrário do triptofano) por mais de um códon. Não é por acaso que no exemplo relatado escolhemos os códons indicados, poderíamos ter codificado o mesmo tripeptídeo com uma sequência de RNAm diferente, como CUC-GCC-CGG-UCC.
Inicialmente, o fato de um único aminoácido corresponder a mais de um tripleto ganhou um significado de aleatoriedade, também expresso na escolha do termo degeneração do código, utilizado para definir o fenômeno da sinonímia. Por outro lado, alguns dados sugerem que a disponibilidade de sinônimos referentes a diferentes estabilidade da informação genética não é de todo acidental, o que parece ser confirmado também pelo achado de um valor diferente da razão A + T / G + C nos diferentes estágios de evolução. Por exemplo, em procariotos, onde a necessidade de variabilidade não é atendida pelas regras do Mendelismo e do neo-Mendelismo, a relação A + T / G + C tende a aumentar, a consequente estabilidade menor, diante das mutações, proporciona maior oportunidades para variabilidade aleatória de mutação genética.
Em eucariotos, em particular em células multicelulares, nas quais é necessário que as células de um único organismo mantenham o mesmo patrimônio hereditário, a relação A + T / G + C no DNA tende a diminuir, diminuindo a oportunidade de mutações em genes somáticos .
A existência de códons sinônimos no código genético levanta o problema, já mencionado, da multiplicidade ou não de anticódons no RNAt.
É certo que existe pelo menos um RNAt para cada aminoácido, mas não é igualmente certo se um único RNAt pode se ligar a um único códon, ou pode reconhecer sinônimos indiferentemente (especialmente quando estes diferem apenas para a terceira base).
Podemos concluir que existem em média três códons sinônimos para cada aminoácido, enquanto os anticódons são pelo menos um e não mais do que três.
Lembrando que os genes são concebidos como trechos únicos de sequências polinucleotídicas muito longas de DNA, é claro que o início e o fim de um único gene devem necessariamente estar contidos na memória.
BIOSSÍNTESE DE PROTEÍNAS
Em diferentes partes do DNA ocorre a abertura da dupla cadeia e a síntese dos diferentes tipos de RNA.
Durante a etapa de carregamento, o RNAt se liga aos aminoácidos (previamente ativados pelo ATP e pela enzima específica). O "maquinário" biossintético é incapaz de "corrigir" tRNAs carregados incorretamente.
O RNAr então se divide em duas subunidades e, ao se ligar às proteínas ribossômicas, dá origem à montagem dos ribossomos.
O RNAm, passando pelo citoplasma, liga-se aos ribossomos, formando o polissoma. Cada ribossomo, fluindo no mensageiro, hospeda gradativamente o RNAt complementar aos códons relativos, pegando os aminoácidos e ligando-os à cadeia polipeptídica em formação.
O RNAt relativamente estável entra novamente na circulação. Os ribossomos também são usados novamente, liberando o polipeptídeo já montado.
O mensageiro, menos estável por ser totalmente monocatenário, é clivado (pela ribonuclease) nos ribonucleotídeos constituintes.
O ciclo continua assim, sintetizando um após o outro os polipeptídeos nos RNAs mensageiros fornecidos pela transcrição.









.jpg)