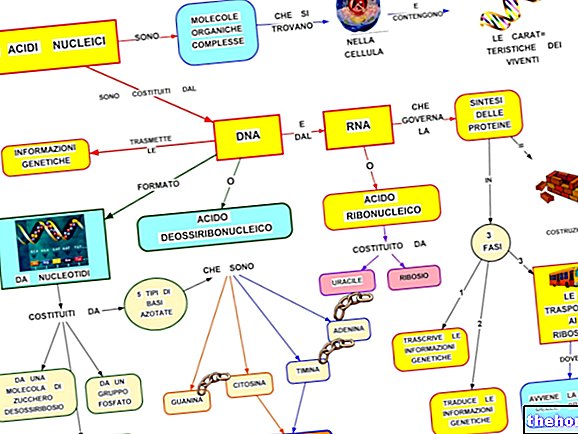
ASSOCIAÇÃO (OU "CONCATENAÇÃO" OU "VINCULAÇÃO")
Até agora, tratamos o cruzamento mendeliano dihíbrido (ou poli-hibrido) assumindo que os diferentes pares alélicos são realmente encontrados em diferentes pares de cromossomos homólogos. Mas o número de pares cromossômicos, embora diferente de espécie para espécie, varia dentro de limites estreitos (poucas espécies atingir cerca de cem cromossomos), enquanto o número de genes pode ser contado na casa das dezenas de milhares.
Que os caracteres escolhidos por Mendel para seus experimentos segregassem todos de forma independente (sem, portanto, confundir os cálculos na distribuição dos fenótipos no F2 do cruzamento poli-hibrido) era uma sorte. Se dois pares de alelos fossem encontrados em loci adjacentes, a lei que que se seguiria seria chamada de lei da associação.
Sabendo que muitos caracteres têm seu locus em um único cromossomo e que eles são os pares cromossômicos que segregam independentemente na meiose, pode-se entender com que freqüência acontece que dois pares de caracteres, se estivessem associados em um cromossomo no parental organismo, permanecerá associado, igualmente, no gameta e, portanto, no organismo para o qual trará seu próprio material genético.
Assim, vemos que a "associação representa uma" exceção, nada rara, à independência enunciada na terceira lei de Mendel.
TROCA OU "CROSSING-OVER" E RECOMBINAÇÃO
Falando em meiose, indicamos que existem dois momentos distintos de mistura do material genético: um é o da segregação dos cromossomos nos gametas, e esse é o observado por Mendel.
O outro momento, que realmente precede, é aquele em que as quatro cromátides de cada par de cromossomos homólogos trocam mutuamente características idênticas. Após essa troca, dois fatores que estavam associados no mesmo cromossomo serão independentes nos gametas. A probabilidade que uma troca ocorra é proporcional, a uma primeira aproximação, ao comprimento do cromossomo, e em cromossomos mais longos pode haver ainda mais do que uma troca.
O fenômeno pode ser detectado citologicamente, observando um número suficiente de meioses sob o microscópio.
A taxa de re-correspondência é a taxa na qual quaisquer dois caracteres que foram associados na geração dos pais se recombinam de maneira diferente em F2.
Se os dois loci forem absolutamente contíguos, a probabilidade de que um quiasma os separe será praticamente nula. A taxa de recombinação será: n ° recombinantes. Se dois loci estiverem em dois cromossomos diferentes, a taxa de recombinação será 0,5 (probabilidade igual, para dois caracteres que foram unidos na geração P, se encontrarem aleatoriamente juntos em F2). A taxa de recombinação pode, portanto, variar entre 0,0 e 0,5. Para pequenas distâncias no cromossomo, a distância e a taxa de recombinação são diretamente proporcionais. Para distâncias maiores, existe a possibilidade de que duas trocas ocorram entre dois loci. Agora parecerá claro que dois fatores separados por duas trocas estão mais uma vez associados. fica claro, neste ponto, que se perde a proporcionalidade entre a distância dos loci e a probabilidade de recombinação.
Os loci encontrados associados no mesmo cromossomo constituem 'grupos de associação'. Loci muito distantes podem ter tal probabilidade de separação por troca que se comportam como independentes, mas cada um deles estará associado, com uma taxa de recombinação mais baixa, aos loci intermediários.
Quando as taxas de recombinação entre muitos pares de genes dentro de um grupo de associação são conhecidas, a construção de 'mapas genéticos' pode começar. Tendo em mente que a distância entre dois genes (aeb) é expressa pela taxa de recombinação e que a distância de a de um terceiro gene c pode ser a soma ou a diferença em relação à sua distância de b, é possível reconstruir um mapa das distâncias recíprocas, que será o mapa genético dentro daquele grupo de associação, ou seja, daquele cromossomo.
Devemos agora considerar em geral alguns conceitos que limitam a manifestação fenotípica de caracteres genotípicos.
Em primeiro lugar, falaremos sobre os conceitos de penetrância e expressividade e, em seguida, dedicaremos atenção especial aos fenômenos de regulação da ação gênica.
PENETRÂNCIA
A penetrância de um gene representa sua capacidade de se manifestar no fenótipo. A penetração é medida estatisticamente contando-se a frequência dos fenótipos que mostram aquele personagem entre 100 genótipos que o contêm. Um traço com penetração de 0,7 é aquele que ocorre fenotipicamente em 70% de sua freqüência genotípica.
EXPRESSIVIDADE
A expressividade é uma avaliação quantitativa do grau de manifestação fenotípica.
REGULAÇÃO DA AÇÃO GÊNICA
As células produzem todas as suas enzimas e proteínas na mesma velocidade e ao mesmo tempo. As células de Escherichia coli, por exemplo, podem receber energia e átomos de carbono do dissacarídeo de lactose, pois são capazes de quebrá-los em glicose e galactose graças à enzima beta-galactosidase. Em uma E. coli normal que pode ter lactose, existem cerca de 3 000 moléculas de beta-galactosidase, o equivalente a 3% das proteínas dessa célula; na ausência de lactose, haverá apenas uma molécula de beta-galactosidase por célula bacteriana. A galactosidase será sintetizada a partir de novas moléculas de mRNA quando puder ser usada. As cepas mutantes de E. coli ricas na enzima são conhecidas mesmo quando a lactose está ausente: esses mutantes são prejudicados em comparação com as células normais, pois são forçados a um consumo desnecessário de energia e materiais para produzir a enzima que permanecerá sem substrato. Substâncias que causam aumento na quantidade de enzimas, como é o caso da lactose, serão chamadas de indutores, enquanto as enzimas serão chamadas de indutíveis. Outras substâncias induzem, também estas de maneira específica, a produção de certas enzimas. Também em E. coli, por exemplo, capaz de construir todos os seus aminoácidos, possuindo carbono e amônio (NH3), a presença no meio de cultura de um determinado aminoácido (histidina, por exemplo) bloqueia a produção de todas as enzimas associadas com a biossíntese do próprio aminoácido: dir-se-á dessas enzimas que são reprimíveis. Nas células bacterianas as moléculas de mRNA são demolidas logo após sua formação, por isso controlar a produção de mRNA significa controlar a síntese enzimática ao mesmo tempo.
O OPERÃO
Para explicar como a célula bacteriana é capaz de controlar sua própria produção de enzimas, Jacob e Monod formularam a hipótese do operon; o operon é formado por vários genes que estão funcionalmente relacionados e alinhados sem descontinuidade ao longo de um trecho de DNA.O operon consiste em três tipos diferentes de genes: o promotor, onde começa a formação do mRNA; o operador, onde o controle é exercido; um ou mais genes estruturais, que codificam para enzimas ou para outras proteínas. No sistema da beta-galactosidase, o operon inclui, além do da beta-galactosidase, também dois outros genes que codificam para a estrutura outras enzimas envolvidas no metabolismo da lactose. Esses genes são adjacentes uns aos outros e são transcritos um após o outro ao longo da mesma hélice de DNA em uma única molécula de mRNA. As moléculas de mRNA assim produzidas são ativas por um período muito curto, após o qual são destruídas por enzimas específicas.
A atividade do operon é por sua vez controlada por outro gene, o regulador, que também pode estar distante do operon: este regulador codifica uma proteína, chamada repressor, que parece se ligar ao DNA no gene operador, sendo o gene operador colocado entre o promotor e os genes estruturais, na verdade, bloqueia a produção de mRNA.
O repressor, por sua vez, é controlado, e o controle é realizado por meio de uma substância "sinal". No caso das enzimas induzíveis esta substância é o "indutor. O indutor" liga-se à molécula repressora modificando sua forma para que ela não possa mais se adaptar ao DNA: neste caso, uma vez que não há repressor entre o promotor e os genes estruturais , o repressor pode formar as moléculas de mRNA e a partir delas as moléculas de proteína. Com o esgotamento do fornecimento do indutor novamente o regulador recuperará o controle, o que interromperá a produção de novo mRNA, portanto de novas proteínas. No sistema da beta-galactosidase o indutor é a lactose ou uma substância muito semelhante a esta. Derivado: eles vai se juntar ao repressor inativando-o de forma a permitir a biossíntese de enzimas. No caso das enzimas repressíveis, a substância que atua como um "sinal" atua como um copressor: o repressor só é ativo se combinado com o co-depressor. No sistema da histidina, que envolve uma dúzia de enzimas diferentes, é esse aminoácido, combinado com seu tRNA, o corepressor, histidina.
INTERAÇÕES ALOSTERICAS
As interações alostéricas, envolvendo a inativação de uma enzima pela alteração de sua forma, fornecem uma maneira diferente de regular a atividade metabólica de uma célula. As interações alostéricas permitem um controle mais preciso do que o sistema indutor-repressor do operon, mas não alcançam o resultado útil de excluir a biossíntese de uma determinada substância do primeiro estágio - a produção de um mRNA.
SISTEMAS DE CONTROLE NO EUCARIUS
Existem alguns fatos que levam a acreditar que um sistema de regulação semelhante ao operon está operando e preeminente entre plantas e animais. Os cromossomos desses organismos diferem profundamente dos de E. coli e de outros procariotos. O controle dos genes nesses as células são muito diferentes. O mecanismo da mitose é tal que cada célula de uma determinada planta ou animal possui todas as informações
genética presente no ovo fertilizado. Portanto, a maioria dos genes em qualquer célula especializada permanecerá ineficiente ao longo da vida da célula. O DNA nessas células está sempre associado a proteínas. Portanto, é possível que a repressão gênica em eucariotos exija exatamente essa associação entre DNA e proteínas.