«Primeiros passos da glicólise
A primeira enzima usada na segunda fase da glicólise é a gliceraldeído 3-fosfato desidrogenase; as desidrogenases são enzimas que catalisam a transferência do poder redutor de uma molécula redutora que se oxida para outra molécula que é reduzida (reação redox). Os substratos dessa enzima são NAD (nicotidamida adenina dinucleotídeo) e FAD (flavina adenina dinucleotídeo).
Nesta etapa, a desidrogenase catalisa a conversão de gliceraldeído 3-fosfato em 1,3-bisfosfoglicerato: no mesmo sítio catalítico, o grupo aldeído é oxidado a carboxila com consequente redução de NAD + a NADH e, posteriormente, o grupo carboxila é capaz para formar uma ligação anidrido com um ortofosfato. O primeiro processo é muito exergônico (libera energia), enquanto o segundo é muito exergônico (requer energia); se não houvesse sítio catalítico, a reação global não ocorreria: a primeira reação ocorreria com a liberação de energia que se dispersaria como calor e que, portanto, não seria aproveitável para formar a ligação anidrido.

Após a formação do 1,3-bisfosfoglicerato, a enzima retoma sua estrutura inicial e está pronta para atuar sobre um novo substrato.

Então vem o fosfoglicerato quinase que permite a transferência de um fosforil de 1,3-bisfosfoglicerato para uma molécula de ADP; obtivemos ATP (um ATP para cada molécula de gliceraldeído 3-fosfato, portanto, dois ATPs para cada molécula de glicose inicial) que compensa o gasto de energia de a primeira fase da glicólise.
O ânion arseniato (AsO43-) afeta a via glicolítica, pois pode substituir o fosfato na primeira reação da segunda fase da glicólise, dando 1-arsênio 3-fosfoglicerato que é altamente instável e, assim que estiver livre do catalisador local, hidrolisa liberando o "arseniato que retorna à circulação. Portanto, o arsenato imita a ação do fosfato e entra no sítio catalítico: na presença do arsenato, a reação que produz ATP (de 1,3-bisfosfoglicerato a 3-fosfoglicerato) não ocorre porque o 3-fosfato gliceraldeído é convertido diretamente em 3-fosfoglicerato; sem ATP disponível, as células morrem (envenenamento por ácido arsênico).
Na terceira reação da fase oxidativa, o 3-fosfoglicerato é convertido em 2-fosfoglicerato pela ação de fosfoglicerato mutase; a reação envolve um intermediário 2,3-bisfosfoglicerato.

Na próxima etapa, uma enzima intervém enolase que é capaz de catalisar a eliminação de uma molécula de água do esqueleto carbonáceo do 2-fosfoglicerato, obtendo-se o fosfoenol pirivado (PEP);

PEP tem um alto potencial de transferência de um fosforil: ele transfere, por meio da ação de uma enzima piruvato quinase, um fosforil a um ADP para dar ATP, na quinta etapa da segunda fase, obtendo piruvato.

O 2-fosfoglicerato e o 3-fosfoglicerato têm um baixo poder de transferência de um fosforil, portanto, para obter ATP a partir dessas moléculas, o 3-fosfoglicerato é convertido em 2-fosfoglicerato, durante a glicólise, porque é obtido desta última. A PEP que é um espécies com alto potencial de transferência.
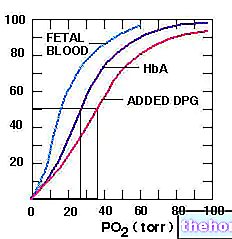
Antes de continuar, vamos abrir um parêntese em 2,3-bisfosfoglicerato; o último está presente em todas as células nas quais a glicólise ocorre em concentração muito baixa (é o intermediário da terceira reação da segunda fase da glicólise). Já nos eritrócitos, o 2,3-bisfosfoglicerato tem concentração estacionária de 4-5 mM (concentração máxima), pois possui um patrimônio enzimático que tem a função de produzi-lo; nos eritrócitos, há um desvio da glicólise para produzir 2,3-bisfosfoglicerato: 1,3-bisfosfoglicerato é convertido em 2,3-bisfosfoglicerato pela ação de bisfosfoglicerato mutase (eritrócito) e 2,3-bisfosfoglicerato, pela ação de bisfosfoglicerato fosfatase (eritrócito) torna-se 3-fosfoglicerato. Em seguida, nos eritrócitos, uma parte do 1,3-bisfosfoglicerato obtido da glicólise é convertido em 2,3-bisfosfoglicerato, que então retorna à via glicolítica como 3-fosfoglicerato; ao fazer isso, a terceira etapa da fase oxidativa de a glicólise da qual o ATP é obtido. A quantidade de ATP perdida é o preço que um eritrócito está disposto a pagar para manter a concentração de 2,3-bisfosfoglicerato de que essas células precisam, porque isso afeta a capacidade da "hemoglobina de se ligar" ao oxigênio.
Vimos que na primeira reação da segunda fase da glicólise o NAD + é reduzido a NADH, mas é necessário que, após a obtenção do piruvato, o NADH seja reconvertido em NAD +: isso ocorre com a fermentação láctica (obtém-se lactato) ou por fermentação alcoólica (piruvato descarboxilase que descarboxila o piruvato e uma desidrogenase que forma etanol entra em ação); fermentações não envolvem oxigênio (anaeróbios).
Devido à fermentação láctica, o ácido láctico, se não for eliminado adequadamente, se acumula nos músculos e, liberando H +, causa contração muscular involuntária e, portanto, cãibras; um músculo em forte estresse também pode atingir um pH mínimo de 6,8.
Por meio do ciclo de Cori, parte da fadiga de um músculo é transferida para o fígado quando o músculo está sobrecarregado. Suponha que o músculo trabalhe sem suprimento de oxigênio (suposição errada): se o músculo trabalhar moderadamente, o ATP necessário para a contração é fornecido exclusivamente pela glicólise. Se a atividade do músculo aumentar e ATP adicional for necessário, acelere o metabolismo aeróbio, convertendo lactato, que é assim eliminado, em glicose. Na realidade, o músculo explora o metabolismo aeróbio: se houver disponibilidade de oxigênio, o músculo explora, sobretudo, o ATP fornecido pelo metabolismo aeróbio e, quando não há mais oxigênio disponível, o metabolismo anaeróbio é acelerado através do ciclo de Cori . Este ciclo pressupõe que o lactato é transferido do músculo para o fígado, onde, ao gastar energia, mais glicose é produzida, que retorna ao músculo. Através deste ciclo, parte do ATP consumido no músculo é fornecido pelo fígado que, através do processo de gliconeogênese, é capaz de produzir glicose que pode ser utilizada pelo músculo para obter ATP.
O metabolismo da glicose descrito até agora não inclui o oxigênio, mas o metabolismo aeróbio da glicose permite obter quantidades 17-18 vezes maiores de ATP do que a obtida com a via glicolítica, portanto, quando a célula tem a possibilidade de escolher entre aeróbio e ed anaeróbio, favorece o primeiro.
No metabolismo aeróbio, o piruvato entra na mitocôndria, onde sofre transformações e, eventualmente, dióxido de carbono e água são obtidos; desta forma, 34 moléculas de ATP são obtidas para cada molécula de glicose degradada.